We will now explore the key details of the AcrB structure that dictate its functions. This segment will highlight the mechanism of drug binding to the monomers of AcrB and eventually elucidate the discovery of a proximal multisite drug-binding pocket in addition to the previously known distal binding pocket. Our aim is to provide a better understanding of the manner in which these drugs are effluxed out of the Gram-negative bacterial cells which express AcrB.
Rifampicin-bound AcrB
Erythromycin-bound AcrB
This essentially proves that large MW drugs such as erythromycin and rifampicin rely on the dynamic drug efflux process, during which they are forced through this channel as the AcrB monomer undergoes transient conformational changes from the access to the binding form. It is the conformational changes and alternating state of the monomers that enable higher MW drugs access to the distal binding pocket in the binding state and eventually be effluxed out via the TolC channels.
The discovery of this proximal binding site in the access monomers hence learly adds to our understanding as to how AcrB has a broad range of specificity.
A Common Pathway
Relevance of Trimeric Structure
- The trimeric crystal structure of AcrB gives us a clue that specific drugs are taken up from the vestibules on the side of the protein and subsequently directed to the TolC channel in the outer membrane before expulsion.
- The efflux transport reaction of the drugs that bind to AcrB can be divided into three intermediate states which are represented by the three monomers of AcrB: Access, Binding and Extrusion.
The Distal-Binding Pocket
- Found in the binding monomer of AcrB.
- Contains the phenylalanine cluster region which is the binding site for low MW drugs.
- Initially perceived as the sole binding site for drugs when the AcrB crystal structure was determined in complex with low MW drugs such as minocyclin (MW ~457) and doxorubicin (MW ~544).
- Further studies of the AcrB structure with a low MW drug bound to its binding monomer revealed intramolecular channels connecting the vestibules to the drug binding site
- However, the exit channel from the binding site through the funnel-like opening at the top of AcrB was found to be closed.
- The neighbouring extrusion monomer meanwhile, had its exit channel opened and a smaller binding site was also found despite it being vacant
- Believed to be the case due to the channel leading from the vestibule to the monomer being closed.
- This structural detail of AcrB when bound to low MW drugs, we believe, led to the conclusion that there exists a functionally rotating mechanism involved within AcrB to transport the drugs out of the cells.
Discovery of a Proximal-Binding Pocket
It was known that AcrB has a wider range of specificity with its drug binding capacity. This lead to further structural studies in search for an explanation that lead to this broad specificity. When the AcrB crystal structure was determined in complex with high MW drugs such as rifampicin (MW ~823) and erythromycin (MW ~734), an additional multisite binding pocket was discovered - the proximal binding pocket. What was more interesting though was that the newfound proximal binding pocket is in the access monomer and NOT the binding monomer, indicating that high MW drugs bind to the access monomer of AcrB.
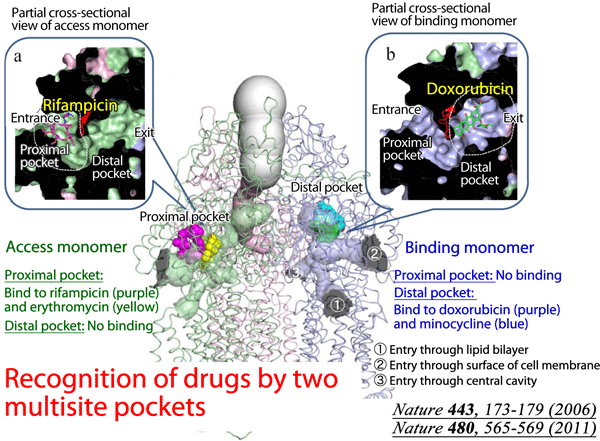 |
Fig 3 - This diagram provides a summary of the binding sites of AcrB and their relative positions within the homotrimer. Note the distinction between access and binding monomers and how they differ in the type of drugs they bind to. The distal binding pocket of the binding monomer is occupied by low MW drugs like doxorubicin and minocycline. The access monomer meanwhile is where the high MW drugs such as rifampicin and erythromycin bind to, specifically to the proximal binding site. |
 |
Fig 4 - When AcrB is crystalised with both rifampicin, a large MW drug and minocyclin, a low MW drug, we find that the drugs bind to different monomers. Rifampicin (in yellow) binds to the proximal binding site of the access monomer (pink) instead of binding directly to the distal binding pocket of the binding monomer (green) as observed with minocycline (in dark blue). |
- Rifampicin, when bound to AcrB, is located between the β-sheets of PC1 & PC2 subdomains within the access monomer. An accurate depiction of the interaction at the molecular level is shown below:
- The Phe-617 residue of the monomer plays a significant role in distinguishing the proximal binding site from the distal binding site
- Phe-617 is described as a hairpin-like loop that is located between the Cβ2' and Cβ3' chains of PC1 subdomain
- When this AcrB structure is comapred to one that is bound with doxorubicin at its binding monomer (distal binding pocket), the Phe-617 loop is found to form a partition dividing the two distinct binding pockets (proximal and distal)
- A narrow channel was also found linking the proximal binding pocket and distal binding pocket, right under the Phe-617 loop
- Phe-617 loop's function becomes distinct here as it was found to swing towards the rifampicin binding site of the binding monomer which directly prevents the binding of rifampicin to the proximal binding site, as observed with lower MW drugs
 |
Fig 5(a) - Rifampicin (in yellow) is sandwiched between the Phe-617 loop (in brown) and the Cβ2' and Cβ3' chains of PC1 subdomain. |
 |
Fig 5(b) - A dissected Pymol generated diagram showing the rifampicin molecule (shown in yellow) forming hydrogen bonds (dotted lines) with the Arg 717 & Asn 719 residues on the β-sheets surrounding the binding site. Atom legend for Arg 717 & Asn 719 residues: Dark Blue - Nitrogen, Red - Oxygen, Light Blue - Carbon. |
Erythromycin-bound AcrB
- Partially overlaps with both the rifampicin and doxorubicin binding sites
- The Phe-617 loop is found to have moved further towards what was the rifampicin binding site
- Believed to be caused by the structure of erythromycin itself- a macrocyclic ring that binds under the Phe-617 loop towards the phenylalanine cluster region
- Also the reason for partial overlap of the erythromycin and doxorubicin binding sites.
- The macrocyclic ring is sandwiched between Phe-617 and a loop containing Ser 134 & Ser 135; Ser residues form hydrogen bonds with the ring
- The distance between the Phe-617 and both Ser 134 & 135 residues is greater in the binding monomer than it is in the access monomer
- This explains why erythromycin does not fit into the the proximal binding site of the binding monomer and instead chooses to bind the proximal binding site of the access monomer
- Optimal geometric complementarity exists between the drug and the proximal binding pocket of the access monomer, enabling interactions (ie. H-bonds) with that greater stabilize binding
- This may also be the reason that erythromycin does not bind to the distal binding site of binding monomer despite it being large enough to accommodate the drug
- Lack of accessibility to the distal binding site may also be caused by the narrow channel under the Phe-617 loop
 |
Fig 6 - The macrocyclic ring of erythromycin (shown in red) is trapped between the Phe-617 loop and the Ser 134 & Ser 135 residues with which it forms a hydrogen bond (dotted lines). Hydrophobic interactions occur between erythromycin and the Phe-617 loop. |
 |
Fig 7 - The images here show the relative position of the Phe-617 loop (in brown) when the proximal binding site of the access monomer is bound to rifampicin (in yellow) and erythromycin (in red) |
The discovery of this proximal binding site in the access monomers hence learly adds to our understanding as to how AcrB has a broad range of specificity.
 |
Fig 8 - The dynamic drug efflux pictured. There are 3 entry channels (2 with known roles) for the high MW drug into the proximal pocket as shown for the access monomer. Conformational change follows to shift the monomer to a binding state which allows high MW drug to move into the distal binding site. The final conformational change leads to the extrusion monomer formation where the drug is effluxed out via the funnel into TolC (not shown in diagram). |
A Common Pathway
An experiment was carried out whereby both high and low MW drugs were added to Gram-negative cultures. It was observed that minocycline, erythromycin and rifampicin inhibited the efflux of doxorubicin and consequently led to its accumulation in cells. This result indicates that low and high MW drugs share a common translocation pathway, indicated by the purple arrows in Figure 8 above.
In the access monomer, both high and low MW drugs can enter the proximal binding pocket (at different times) via either one of the entrance channels. One channel leads from the vestibules and the other from the grooves, as mentioned in the subtopic "Proposed Transport Mechanisms" under Structure. High MW drugs bind to the proximal pocket but the smaller low MW drugs are directed past it and end up binding the distal pocket in the binding monomer. In the extrusion monomer, both types of drugs are channelled up and out of AcrB via a funnel, toward TolC. The 2 entrance channels, 2 binding pockets and TolC funnel thus constitute a common intramolecular pathway for efflux.
In the access monomer, both high and low MW drugs can enter the proximal binding pocket (at different times) via either one of the entrance channels. One channel leads from the vestibules and the other from the grooves, as mentioned in the subtopic "Proposed Transport Mechanisms" under Structure. High MW drugs bind to the proximal pocket but the smaller low MW drugs are directed past it and end up binding the distal pocket in the binding monomer. In the extrusion monomer, both types of drugs are channelled up and out of AcrB via a funnel, toward TolC. The 2 entrance channels, 2 binding pockets and TolC funnel thus constitute a common intramolecular pathway for efflux.
I never knew that efflux proteins had such complex mechanisms involving proximal and distal sites. Always thought it was just a plain ol' channel!
ReplyDeleteTry another colour for the drugs in Fig 6 and 7? Maybe something lighter would make it pop out more. Fig 3 could use more explanation.
ReplyDeleteCheck out my blog at http://qscreceptor.blogspot.co.uk/
Good diagrams. Clearly layed out and good outside reading.
ReplyDelete